私たちが普段摂取する食物のエネルギーを、ATPという体内で利用可能なエネルギー源に変換してくれる機能を担っているのが、ミトコンドリアです。
ミトコンドリアではまず、クエン酸回路という代謝経路を経てNADHやFADH2が作られます。さらにそのNADHやFADH2を使って多数のATPを産生します。この過程を電子伝達系と言います。
今回はその電子伝達系について詳しく解説していきます。
電子伝達鎖の概要
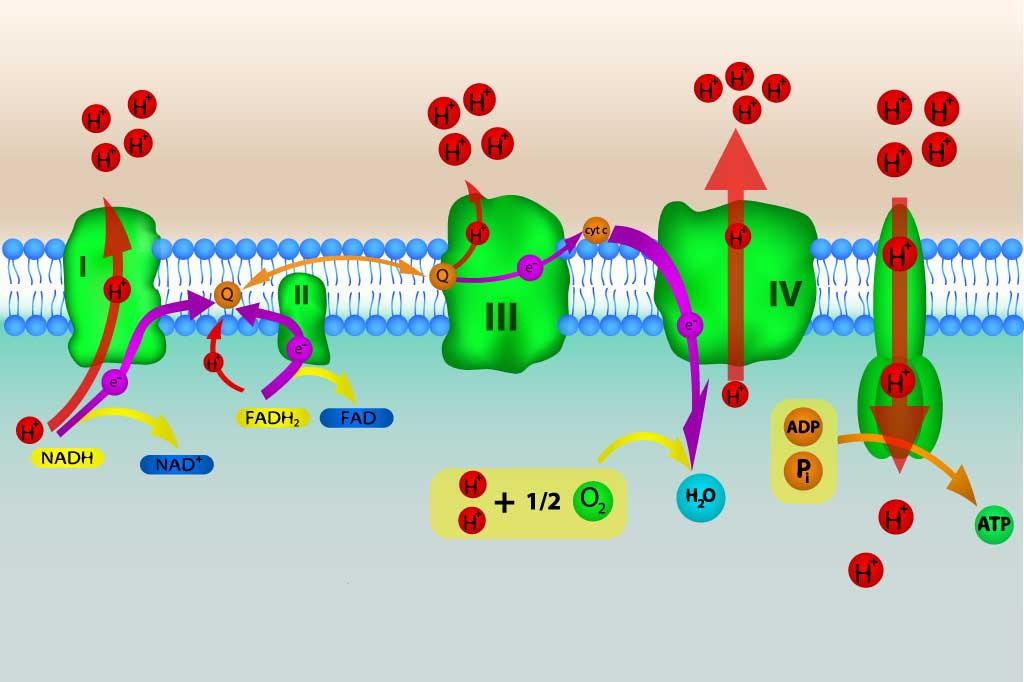
ミトコンドリアのマトリックス膜には3つの酵素複合体が埋め込まれていて、これら3つの複合体内を電子が移動するとH+がマトリックス内から膜間腔へと汲み出されます。
このように、電子を伝達する複合体群を総称して電子伝達鎖と呼びます。特にミトコンドリアの電子伝達鎖は食物からエネルギーを生産する呼吸の過程でもあるため、呼吸鎖とも呼ばれます。
これら3つの複合体は複合体Ⅰ・複合体Ⅲ・複合体Ⅳといって、それぞれNADH脱水素酵素複合体、シトクロムc還元酵素(別名:シトクロムb-c1複合体)、シトクロム酸化酵素複合体とも呼ばれます。
複合体Ⅳ(コハク酸脱水素酵素)も実はあって、これについてはあとで解説します。

呼吸鎖の機能はまず、複合体ⅠがNADHから電子を受け取ってユビキノンに渡します。ユビキノンは電子を受け取るとユビキノールという物質に変化します。
ユビキノールはさらに電子を複合体Ⅲに渡し、複合体Ⅲも受け取った電子をシトクロムcに渡します。最後にシトクロムcから複合体Ⅳに電子が渡され、酸素と反応して水が生成することで電子伝達が終了します。
この間、電子が通るのに合わせて3つの複合体はH+をマトリックスから膜間腔へ汲み出します。
するとマトリックスと膜間腔のあいだにH+の濃度勾配が形成され、それを解消しようとATP合成酵素内をH+が通り抜けることでマトリックスに戻ります。
このときH+がATP合成酵素内を通り抜けたことで中のダービンが回転し、ADPをATPへと変換します。これを酸化的リン酸化と呼びます。
さて複合体Ⅱについてですが、これは複合体ⅠとⅡの間に位置していて、FADH2から電子を受け取ってユビキノンに渡します。他の複合体と違うところは、電子伝達の際にH+をマトリックスから膜間腔へ汲み出さないことです。
ということで、各複合体についてもう少し詳しく見ていきましょう。
NADH脱水素酵素(複合体Ⅰ)
NADHA脱水素複合体は膜内タンパクと膜外タンパクによって構成され、その分子量は100万にものぼります。複合体Ⅰは電子を伝達する領域とH+を汲み出す領域に分かれています。
まず、複合体Ⅰ内のフラビンモノヌクレオチド(FMN)が、NADHの酸化によって生じた電子を受け取ります。FMNは酸化還元酵素の補欠分子族です。
NADH → NAD+ + H+ + 2e–
電子は8つの鉄-硫黄クラスターを介してユビキノンに伝達されます。電子が2つ(NADH1個分)ユビキノンに伝達されるごとに4個のH+がマトリックスから膜間腔へ汲み出されると考えられています。
鉄-硫黄タンパクと呼ばれるタンパク質は活性中心に2~4個の鉄をもち、硫黄原子とシステイン残基の側鎖に結合して鉄-硫黄クラスターを形成しています。1回で電子1個を運ぶことができます。

シトクロムc還元酵素(複合体Ⅲ)
ユビキノン(ubiquinone)は、キノン(quinone; Q)や補酵素 Q(coenzyme Q)とも呼ばれます。コエンザイムQ10は聞いたことがあるのではないでしょうか。サプリメントとして売られているものです。
さて、ユビキノンは電子を2個まで授受することができ、1個受け取るごとにH+を1つH2Oから奪って構造が変化します。1個電子を受け取るとユビセミキノン(QH•)になり、もう1個受け取るとユビキノール(QH2)になります。
NADH1つからは電子が2個放出されるので、ユビキノンがちょうど1つユビキノールになる感じです。
呼吸鎖内では、ユビキノールは複合体Ⅰから複合体Ⅱへと電子を運びます。ユビキノールからの複合体Ⅲへの電子の受け渡しは2段階に分かれていて、Qサイクルと呼ばれています。
- ユビキノール(QH2)が電子2個を複合体Ⅲに渡しと同時に、H+を2つ膜間腔に放出する。渡された電子のうち1個はシトクロムcに伝達され、もう1個は別のユビキノン(Q)に戻されてユビセミキノン(QH•)になる。
- 再び別のユビキノールが(QH2)が電子を2個複合体Ⅲに渡してH+を2つ膜間腔に放出する。渡された電子のうち1個はシトクロムcに伝達され、もう1個はユビセミキノン(QH•)に渡されてユビキノール(QH2)になる。
このようにして、ユビキノールが渡す2つの電子のうち、1つはシトクロムcに伝達され、もう1つはキノンに戻されてプールされます。結果的にシトクロムcに電子が2個渡されることでH+が合計4つ膜間腔に放出されることになります。

シトクロムc酸化酵素(複合体Ⅳ)
シトクロムcに電子が渡され、最後に行き着く複合体がシトクロムc酸化酵素です。シトクロムcからは電子が1個ずつ渡され、酵素複合体内の銅イオン、ヘムaを経てヘムa3へと渡ります。
ヘムa3には銅イオンとの間にO2分子が捉えられていて、電子が4個たまると下の酸化反応を起こします。この酸化反応が1回起こるごとにH+が4つマトリックスから膜間腔へと汲み出されるのです。電子が酸素と反応して水になることで電子伝達が終了します。
O2 + 4H+ + 4e– → 2H2O
このようにして、NADHから放出された電子が呼吸鎖内を移動することでH+がマトリックスから膜間腔へと汲み出されることでATP合成の準備が整うのです。

ATP合成酵素
電子伝達によってできたマトリックスと膜間腔の間のH+濃度勾配の解消は、ATP合成と共役して起こります。
濃度勾配を解消しようと、H+はATP合成酵素(F1F0-ATPアーゼ)の分子内を通ってマトリックスに流入します。このATP合成酵素は23以上のタンパクサブユニットにより構成され、総分子量は60万になります。
H+の電気化学的な勾配にしたがってADPからATPを合成することができます。逆に、ATPを加水分解することで電気化学的な勾配を作ることもできるのです。
ATP合成酵素は回転子と呼ばれる回転部分と、それを固定する固定子に分かれています。H+はまず膜間腔側からATP合成酵素のaサブユニット内に入ってcサブユニットに結合し、最終的にaサブユニットからマトリックス側に流入します。
H+がcサブユニットに結合するとc・ε・γサブユニットが回転します。γサブユニットが回転するとα3β3サブユニットの立体構造が変化していき、それに伴ってADPからATPが合成されます。










