“遺伝子が発現する”という表現を耳にしたことがあるかもしれません。
これはDNAの塩基配列に書き込まれた遺伝情報、つまりタンパク質の設計図をもとに実際にそのタンパク質が出来上がることを意味しています。
ここでは遺伝情報からタンパクが生成するまでの過程を簡単に見ていきましょう。
目次
セントラルドグマとは
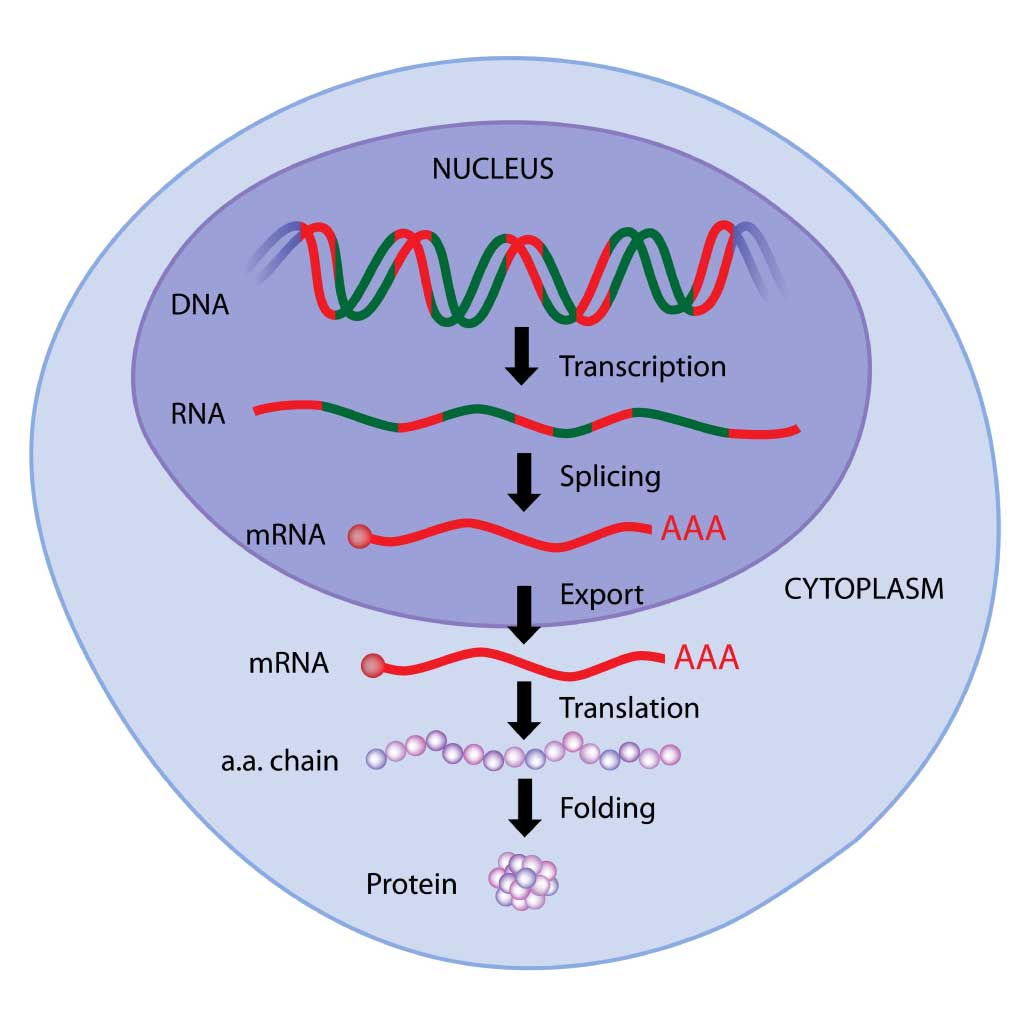
私たちの遺伝情報はDNAに塩基配列として書き込まれていますが、その塩基配列は読み取られ、RNAと呼ばれる写しが出来ます。この過程を転写といいます。
さらに出来上がったRNAもその塩基配列が読み取られることでタンパク質が出来上がります。この過程を翻訳といいます。そしてこの出来上がったタンパク質が体の中で様々な役割を果たすのです。
DNAは細胞分裂する際にコピーされることで、分裂した後の細胞でも保存されます。この過程を複製といいます。
このような生物の遺伝子に関する基本法則のことを、分子生物学者フランシス・クリックはセントラルドグマと提唱しました。次の項からはセントラルドグマの複製・転写・翻訳についてもう少し具体的に見ていきます。

セントラルドグマの図 実線の矢印が複製・転写・翻訳に対応しています。破線の矢印は特別な場合の情報伝達を示しています。
転写
DNAの塩基配列を読み取ってRNAを合成する酵素をRNAポリメラーゼといいます。酵素の主成分はタンパク質であるため、このRNAポリメラーゼもまたDNAの転写・翻訳によって生成したものです。

転写バブルの図 DNAの二重らせんの一部がほどけて転写バブルを形成。二本鎖DNAのうち鋳型鎖とワトソン・クリック塩基対が作れるようにRNAの3’末端にヌクレオチドが付加されていく。
二本鎖DNAであれば二本のうち一本のみを転写の型(鋳型鎖といいます)として3’→5’方向に塩基配列を読み取りながら、鋳型鎖とワトソン・クリック塩基対が作れるようにヌクレオチドを選んで、伸長するRNA鎖の3’部位につないでいきます。
DNA鎖には転写を調節する配列が存在し、RNAの転写を開始する部位と転写の速度を決めています。この転写調節部位はふつう転写開始部位よりも鋳型鎖の3’側の上流に位置しています。
原核生物の場合はアクチベーターとリプレッサーと呼ばれるタンパク分子が転写調節部位に結合して転写開始の促進と抑制を行います。真核生物であれば転写因子がその機能を果たします。
転写後修飾
ほとんどの原核生物において、転写によってできたRNAはそのまま塩基配列が読み取られて翻訳され、目的のタンパク質が出来上がります。
一方で真核生物の場合、転写によってできたばかりのRNAは翻訳される前に修飾されます。5’末端と3’末端にはそれぞれ”キャップ”構造と”ポリA尾部”構造が形成され、スプライシングという作業に入ります。
スプライシングではRNAが部分的に切り取られて、残った部分が元の順序のまま再びつながります。このときスプライシング部位を変えることで最終的なRNAの配列に違いが生まれ、同じDNAの塩基配列から別のタンパク質を合成することができます。
このように、修飾前のRNAを一次転写物、修飾後のRNAを成熟RNAと呼んだりします。

スプライシングの図 mRNAは転写開始後まもなく5’キャップ構造が形成される。ポリA尾部は転写が終了してから修飾され、スプライシングが始まる。切り取られる領域をイントロン、残りの領域をエキソンという。
翻訳
いよいよ、転写によって出来上がったRNAの情報を読み取ってタンパク質を作っていきます。
実はRNAの中には翻訳されないものもあり、それらと区別するためにmRNA(messenger RNA)という言い方をします。DNAに記されている情報を伝達するという意味でmessengerという意味が使われているわけです。
ポリペプチド鎖の合成
翻訳ではリボソームと呼ばれるRNAとタンパクの複合体がmRNAに結合し、アミノ酸を順につなぐことでタンパク質を合成します。RNA中の3つの連続する塩基が、20種類のアミノ酸の中から1つを指定します。
この各アミノ酸に対応する塩基3つの配列のことをコドンと呼びます。このコドンに従ってリボソーム-mRNA複合体にアミノ酸を運んでくるのがtRNA(transfer RNA)です。アミノ酸の運び屋という意味でtransferですね。

tRNAとmRNAの対応図 コドンが指定するアミノ酸をコドンに相補的なアンチコドンを持つtRNAが連れてくることでポリペプチド鎖が伸長する。AUGは開始コドンであると同時にメチオニンを指定する。
tRNAはアンチコドンと呼ばれるコドンに相補的な配列を持っていて、対応するアミノ酸が結合してアミノアシルtRNAを形成するとmRNAに結合してリボソームが運んできたアミノ酸を伸長中のアミノ酸のC末端につなげます。

翻訳の図 リボソームを介して、ポリペプチド鎖のC末端に運ばれてきたアミノ酸が順に付加していく。実際には1本のmRNAに対して複数のリボソームが時間差で翻訳を行なっていく。
こうしてポリペプチド鎖が完成し、その後の正しい折りたたみや高次構造の形成によってタンパク質が出来上がります。タンパク質は消化酵素や触媒酵素、筋肉、物質の貯蔵・運搬などの役割を果たしていきます。
遺伝暗号
mRNAの塩基の種類はA,U,G,Cの4つでRNA中の3つの連続する塩基が1つのアミノ酸を指定します。
従って、考えられるコドンは全部で43=64種類です。
標準アミノ酸は全部で20種類であるため、異なるコドンが同一のアミノ酸を指定することもあります。
ただし、コドンはアミノ酸以外にも翻訳の開始と終結も指定する必要があります。これらのコドンはすでに分かっていて、開始コドンはAUGでメチオニンも同時に指定します。終始コドンは、UAA,UAG,UGAの3種類です。
複製
最後にDNAの複製を見て終わりにしましょう。
DNAを複製、つまりコピーして数を2倍にするためにはまず二本鎖DNAの二重らせんをほどいていきます。
ほどかれた二本のDNA鎖(親鎖)は同時に鋳型にされて相補的な塩基対が伸長していきます。
鋳型鎖と新しく伸長してできたDNA鎖(娘鎖)がそのまま二重らせんを形成することで複製が完了し、2つの二本鎖DNAが出来上がります。
このとき、どちらの二本鎖DNAにも元のDNAが半分ずつ継承されているため、この複製を半保存的であるといいます。
DNAの新規合成
さて、新しいDNA鎖を伸長するのがDNAポリメラーゼなのですが、RNAポリメラーゼと違ってDNAの新規合成ができないため既存のDNA鎖を伸ばすことしかできません。
そこで一番最初の伸長開始から数塩基分はRNAポリメラーゼの1種であるプライマーゼがRNAを合成し、そこからDNAポリメラーゼがDNAとして伸長していきます。このRNAの部分をプライマーと呼びます。

複製フォークの図 DNAポリメラーゼは5’→3’の方向にしか合成することができない。そのため2つの娘鎖のうち一方はリーディング鎖として連続的に合成されるが、もう一方はラギング鎖として不連続に合成される。
合成された娘鎖はこのままだとRNAを部分的に含んでしまうため、複製の最後にRNAの部分のみを刈り取ってDNAに置き換える作業をします。
伸長の段階に話を戻しますが、DNAポリメラーゼは既存のDNA鎖を5’→3’方向にしか伸ばすことができません。一方の親鎖は3’→5’方向に伸びているため娘鎖は5’→3’方向に連続的にコピーしていくことができます。これをリーディング鎖(先行鎖)といいます。
もう一方の親鎖は5’→3’方向に伸びているため娘鎖を連続的に伸長しようとすると3’→5’方向になってしまうため不可能です。そこでまず、プライマーゼがRNA鎖を部分的に合成し、RNAの断片同士を5’→3’方向にDNAポリメラーゼが伸ばしてつなげていくことで娘鎖を伸長させます。
このようにして伸長していくDNA鎖をラギング鎖(遅行鎖)といいます。ラギング鎖を合成するためにはRNA断片がその都度必要になり、この断片を発見した分子生物学者岡崎令治にちなんで岡崎フグメントと呼ばれています。
RNAプライマーをDNA配列へ
先ほども言ったように、複製を終わらせるためにはRNAプライマーをDNAに正しく置き替えなければなりません。
例えば大腸菌には3種類のDNAポリメラーゼがあって、DNAポリメラーゼⅢ(PolⅢ)はリーディング鎖とラギング鎖の大部分を伸長します。
一方、DNAポリメラーゼⅠ(PolⅠ)はヌクレオチド鎖の末端から数塩基を加水分解して刈り取る5’→3’エキソヌクレアーゼ活性を持っています。PolⅠはRNAプライマーを刈り取って空いたところを5’側から伸長し直し、最後にDNAリガーゼが切れ目をつなぎます。

DNA配列の校正
ところでRNAポリメラーゼは約104塩基につき1回の割合で塩基を間違えて合成しますが、DNAポリメラーゼは108~109塩基につき1回の割合でしか塩基を間違えて合成しません。
これはDNAの複製中にワトソン・クリック塩基対ではない塩基対が形成された際にそれを検出して校正する機能がpolⅠとPolⅢの両方に備わっているからです。
この機能は3’→5’エキソヌクレアーゼ活性といって、伸長中のDNA鎖の3’側から5’方向に数塩基刈り取ります。これ以外にもDNAの損傷を検出し修復する酵素群が備わっています。








