植物は普段、私たち人間と違って光エネルギーを利用して二酸化炭素と水から有機物を生成しています。この働きを光合成といって、細胞小器官の1つである葉緑体のなかで行われているのでした。
今回は葉緑体について、その構造や光合成の反応の詳細を解説していきたいと思います。
目次
構造
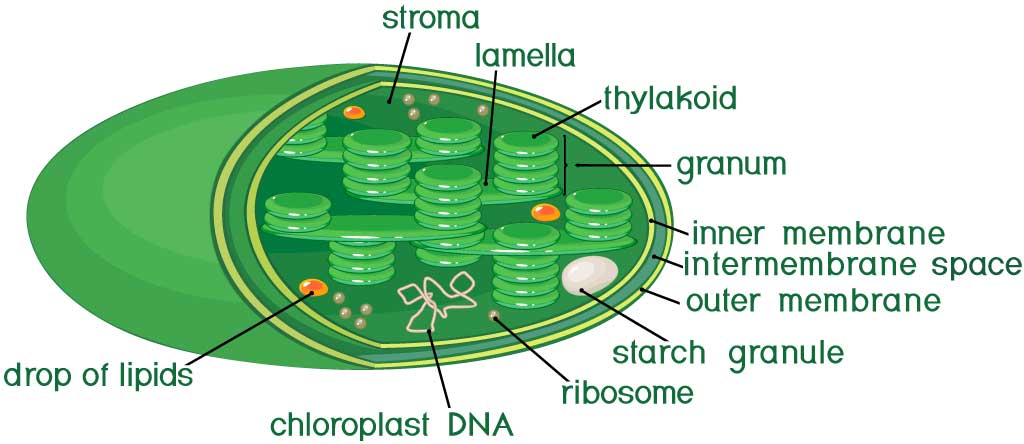
何かとミトコンドリアと比較される葉緑体ですが、主な共通している点といえば外膜と内膜、独自のDNAを持っていることでしょう。したがって、独自のリボソームや葉緑体DNAから転写されたRNAが存在します。
これら遺伝系は内膜内のストロマと呼ばれる領域にあります。ミトコンドリアで言うところのマトリックスですね。ただし、葉緑体にはミトコンドリア内膜の陥入構造のようなクリステが存在しません。
その代わりにチラコイドという扁平で円盤状の膜構造がストロマの内部にあります。さらに、チラコイド膜が折りたたまれて積み重なったグラナという構造も形成しています。ラメラもチラコイド膜による構造体で、異なるグラナ同士をつないでいます。

光合成の概要(明反応と暗反応・光合成電子伝達と炭素固定)
中学校や高校で習う光合成の反応式といえばこんなものでしょうか。
6CO2 + 12H2O + 光エネルギー → C6H12O6 + 6H2O + 6O2
もしくは両辺から6H2Oを引いて、
6CO2 + 6H2O + 光エネルギー → C6H12O6 + 6O2
実際には光合成により水も生成するので1つ目の式の方がより正確ですね。反応の過程をもう少し詳しく見てみましょう。光合成の反応過程は主に2つの段階に分けることができます。

step1. 光合成電子伝達反応(明反応)
葉緑体のチラコイド膜には、ミトコンドリアのクリステと同じように電子伝達を行うことのできる酵素複合体が並んでいます。ここに光が当たると中にあるクロロフィルから電子が飛び出して(光化学反応)、電子伝達鎖内を移動します。
この電子伝達過程でH+がストロマからチラコイド内腔へ汲み出されます。形成されたH+の濃度勾配を解消しようとATP合成酵素内を通り抜け、ストロマに戻るのと共役してADPからATPが合成されます(光リン酸化)。
最後にNADP+が電子(とH+)を受け取ってNADPHになることで電子伝達反応は終了します。
グルコース1分子の生成を想定すると、この過程でH2Oが12分子消費され、O2が6分子生成します。
step2. 炭素固定反応(暗反応)
step1で生成したATPとNADPHは、カルビン・ベンソン回路と呼ばれる代謝経路でエネルギーとして利用されます。ここではCO23分子の炭素から最終的にグリセルアルデヒド3-リン酸を合成します。
グリセルアルデヒド3-リン酸はその後、スクロースやデンプンに変換されたり、解糖系に組み込まれてATP合成にエネルギー源になったりします。
グルコース1分子の生成を想定すると、この過程でCO2が6分子消費され、H2Oが6分子生成します。
光化学系
葉緑体のチラコイド膜に膜タンパクとして埋め込まれている電子伝達鎖のなかには、光化学系と呼ばれる酵素複合体があります。
これら光化学系には、ミトコンドリアの電子伝達酵素複合体と同じく電子伝達のためのヘムや鉄-硫黄クラスター・キノンが含まれているだけでなく、多数のクロロフィルも結合しています。
クロロフィルは植物の色をなす緑色の色素で、疎水性の尾部とマグネシウム原子を中心にもつポルフィリン環より成り立っています。
クロロフィルが太陽光からの光子を1個吸収すると、クロロフィル内の電子の1つが励起されてエネルギーの高い状態になります。励起された電子は元の基底状態に戻ろうとするため以下のうち、いずれかの反応が起きます。
- 分子の熱運動や蛍光を発することによってエネルギーを放出する。
- 励起した電子のエネルギーのみを隣のクロロフィルに渡す(共鳴エネルギー移動)。
- 励起した電子を別の分子に渡し(電荷分離反応)、基底状態の電子を別の分子から奪い取って元に戻る。
光化学系に含まれているクロロフィルの場合、2番目と3番目の反応が起きます。ほとんどの場合、励起した電子のエネルギーのみを隣のクロロフィルに渡して基底状態に戻るわけですが、それでは電子伝達は起こりません。
スペシャルペア
特別な場所に位置するクロロフィルのペアのみが、電荷分離反応を起こすことができるのです。このペアをスペシャルペアと呼びます。
スペシャルペアに光が当たると、光子エネルギーによって一方のクロロフィルの電子が励起されるとともにペアが解離されます。
すると励起した電子を隣のクロロフィルに渡すことができなくなるため、電子は同じ複合体中を移動してプラストキノンに渡されます。
このようなスペシャルペアをもつ複合体を光化学反応中心と呼びます。スペシャルペアではないクロロフィルを含む複合体をアンテナ複合体と呼び、これら2種類の複合体が光化学系を構成しています。
アンテナ複合体
光合成を行うにあたって、スペシャルペアが太陽光から光子を直接受け取るだけでは反応が遅く、効率も悪いです。
効率よく光合成を行うためには1秒間に数百個の光子を吸収する必要がありますが、スペシャルペアは1つにつき1秒間で光子を約1個しか吸収することができません。
そこでアンテナ複合体中のクロロフィルが太陽光から吸収した光子エネルギーを、共鳴エネルギー移動によってスペシャルペアに送ってくれます。
これによってスペシャルペアは単位時間当たりに十分なエネルギーを吸収して電子を励起させることができるのです。
ゼットスキーム
さて、電子伝達鎖に光が当たってからATPとNADPHを合成するまでの過程をもう少し詳しく見ていきます。太陽光によって励起されたエネルギーは以下のようにして電子伝達鎖内を移動し、最終的にNAD+にたどり着きます。

さらに、光合成の際に電子伝達を行う分子の酸化還元電位の変遷を見ると下図のようになります。その変遷を90°回転させるとZの形に見えますよね。このことからゼットスキーム(Z sheme)なんて呼ばれたりもします。

酸化還元電位が高い物質ほど電子を保持しやすく、低い物質ほど手放しやすいと考えておけば良いです。
それでは各電子伝達体について詳しく見ていきましょう。
光化学系Ⅱ
上の光合成の概要図を見ればわかるように、実は光化学系には光化学系Ⅰと光化学系Ⅱの2種類があります。まず光エネルギーを利用して伝達する電子を用意するのが光化学系Ⅱです。
順番的に光化学系Ⅰが最初にくるのではと思うかもしれませんが、1番目に発見されたから光化学系Ⅰ、2番目に発見されたから光化学系Ⅱと名付けられています。
光化学系Ⅱに含まれているスペシャルペアが光子を吸収したり、アンテナ複合体からの共鳴エネルギー移動によってエネルギーを吸収すると、一方のクロロフィルの電子が1つ励起されてプラストキノンに渡されます。
これによって電子伝達が始まるのです。
電子をプラストキノンに渡せばそれで終わりというわけではありません。一方のクロロフィルは電子を1つ失ったわけですから、電子をどこか別の分子から奪って元に戻る必要があります。その奪う相手というのが水です。
普段私たちが摂取している水は当然安定なはずですから、電子を引き抜くのもそう簡単なはずがありません。現に水の標準酸化還元電位は+820 mvとかなり高いです。
酸化還元電位が高いということは、電子を受け取りやすく手放そうとしないことを意味しています。従って、これより酸化還元電位の高い物質を用意しないと自ら電子を引き抜くことはできません。
それを実現するのが光化学系Ⅱに含まれるスペシャルペアのクロロフィルです。このペアは光の吸収波長を680 nmにピークをもつことから、P680といわれています。その酸化還元電位は+1270 mvと非常に強力です。
結果、光化学系Ⅱでは以下の反応が起きます。
2H2O + 4つの光子 → 4H+ + 4e– + O2
生じたH+はチラコイド内腔に流入することで濃度勾配形成に利用されます。
シトクロムb6-f複合体
プラストユビキノンは電子を2つ受け取るとプラストユビキノールになります。すると光化学系Ⅱの隣に位置するシトクロムb6-f複合体まで電子を運びます。
ミトコンドリアの複合体Ⅲとユビキノールの関係のように、Qサイクルによってプラストユビキノールから放出された2個の電子のうち1つはプラストシアニンに渡り、もう1つは再びキノンに戻ります。
このとき生じたH+も、ミトコンドリアの場合と同じようにチラコイド内腔に流入することで濃度勾配が形成されます。

光化学系Ⅰ
電子を受け取ったプラストシアニンは光化学系Ⅰへと移動すると、光化学系Ⅰに含まれるスペシャルペアに電子を渡します。このスペシャルペアは700 nmの波長の光を吸収するためP700と呼ばれています。
P700はさらに太陽光の光子を吸収することで電荷分離反応を起こし、電子を光化学系内の別の分子に渡します。
電子は光化学系Ⅰ内を移動し、クロロフィルA0に到達するとさらに2個のプラストキノン、3個の鉄-硫黄クラスターを経てフェレドキシンへと運搬されます。
フェレドキシンは受け取った電子をチラコイド膜に結合しているフェレドキシン-NADP+還元酵素(FNR)に渡し、FNRは電子を使ってNADP+からNADPHを合成します。
また、電子伝達の際にチラコイド内腔とストロマの間にH+の濃度勾配が形成されます。この濃度勾配を解消しようとH+がストロマに流入するのと共役して、チラコイド膜に埋め込まれているATP合成酵素がADPからATPを合成します。この過程はミトコンドリアのATP合成と似ていますね。









